Interneuron Function and Dysfunction
Interneuron development is intimately linked to their ultimate function. We have demonstrated how perturbation of the developmental programs utilized by specific interneuron populations affect their development in ways that may manifest in disease. For instance, Mef2c and Satb1 are required for the development of PV and SST interneurons (Mayer et al. Nature 2018, Allaway et al. Nature 2021). To explore the role of interneurons in circuit assembly, the Fishell laboratory has for some years studied how sensory inputs shape the formation of cortical inhibitory circuits (Tuncdemir et al., Neuron 2016). In a recent study we demonstrated how bottom-up signaling to interneurons in layer 1 is required for the maturation of top-down circuits (Ibrahim et al., 2021). Moreover, we have identified that synaptic pruning of inhibitory synapses is a selective and specialized process. We showed that GABA-receptive microglia selectively interact with inhibitory cortical synapses during a critical window of mouse postnatal development.(Favuzzi et al., Cell, 2021). In addition to revealing events critical to normal development, improved understanding of how cortical circuits are constructed may provide insight into the etiology of psychiatric disorders. Finally, a new direction of the laboratory involves the systematic search for enhancers. These can be used in the context of viruses to target specific interneuron populations (see topic 4). This approach provides unprecedented opportunity for exploring cortical development and function in both mice, as well as less amenable species including non-human primates.
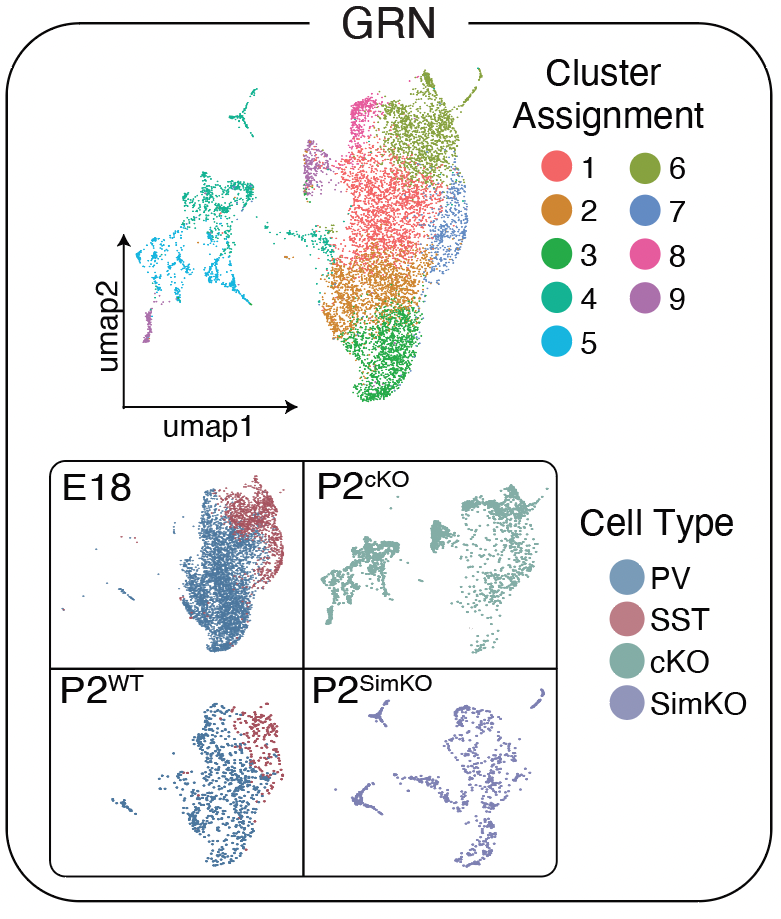
Figure 1 - Predicting Mef2c gene changes in interneurons in silico
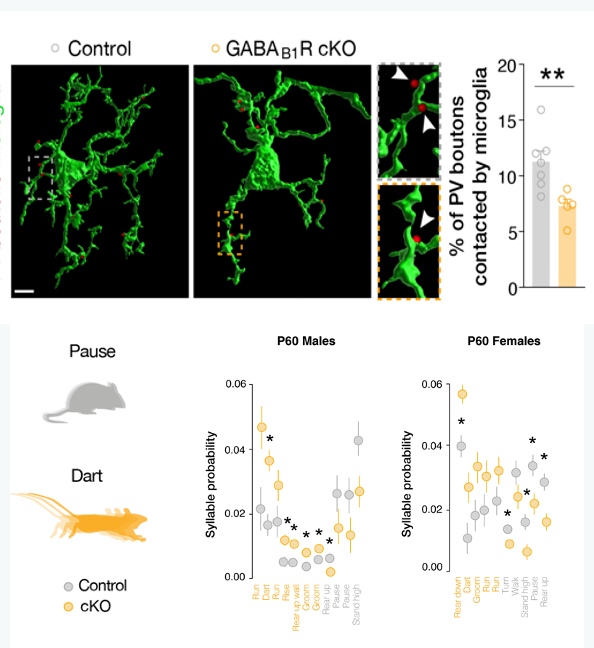
Figure 2 - Loss of GABAB1R in microglia prevents inhibitory synapse pruning and affects gender-specific behavior

Figure 3 - Targeting SST interneurons in Macaque with enhancer-mediated AAV
Selected References
Tuncdemir S, Wamsley B, Stam FJ, Osakada F, Goulding M, Callaway EM, Rudy B, Fishell G. Early somatostatin interneuron connectivity mediates the maturation of deep layer cortical circuits. Neuron 2016; 89:521-535.
Mayer C, Fishell G. Developing neurons are innately inclined to learn on the job. Nature. 2018 Jul 23; doi 10.1038/d41586-018-05737-2.
Allaway KC, Gabitto MI, Wapinski O, Saldi G, Wang C-Y, Bandler RC, Wu SJ, Bonneau R, Fishell G. Genetic and epigenetic coordination of cortical interneuron development. Nature. 2021 Sep;597(7878):693–697. PMID: 3455224
Favuzzi E, Huang S, Saldi GA, Binan L, Ibrahim LA, Fernández-Otero M, Cao Y, Zeine A, Sefah A, Zheng K, Xu Q, Khlestova E, Farhi SL, Bonneau R, Datta SR, Stevens B, Fishell G. GABA-receptive microglia selectively sculpt developing inhibitory circuits. Cell. 2021 Jul 22;184(15):4048-4063.e32. PMID: 34233165